Яков Гельфандбейн - Статьи
Константы карциногенеза - тайна метаморфозы Эшера
"Одна из поразительных
особенностей природы - многообразие возможных схем ее толкования..."
Richard Feynman
Ситуации и отношения на клеточных множествах.
|
Уважаемый читатель! Позволю себе напомнить, что нас интересуют проблемы возникновения и развития злокачественных опухолей. Слов нет - тема весьма неприятна. Но ее невозможно обойти вниманием. За человеческую жизнь надо бороться. И бороться, вооружаясь самыми всесторонними и разнообразными знаниями, не жалея сил, запасшись терпением. И быть готовым ко всяким сложностям. И в этой проблеме мы хотим узнать закономерности изменения картины, которую мы наблюдаем под микроскопом в срезах биологического субстрата. Именно эта картина, содержит и отражает всю информацию о процессе! Но сложность ее познавания связана с тем, что мы никогда на можем видеть этот процесс с начала до конца в принципе: ведь в срезе - множество клеток, изъятых из организма. Их вид "заморожен". И в других срезах - то же самое, но к тому же, это ведь уже и другие клетки! А мы хотим знать не только то что происходит с их множествами - во времени, в ходе всего процесса и в его пространстве, но и с каждой отдельно взятой клеткой. Даже то, что произошло бы с той клеткой, которая была изъята, если бы этого не было сделано! Нас интересует процесс! Вот в чем сложность! Задачу можно сравнить разве только с неким гибридом ребуса и кроссворда, построенным на неизвестном нам языке! Чтобы понять с чем мы имеем дело, необходим с одной стороны инструмент для разъяснения, а с другой - объект, схожий с процессом, на котором можно опосредовать эти разъяснения. Эшер дал нам такую возможность. Объект нашего познания чрезвычайно сложен, проблема его познания поставлена с позиций, необычных даже для специалиста, не говоря хотя бы просто о любознательном читателе. В такой постановке задача исследования клеточных полей ранее не ставилась вообще. Именно серьезность и сложность излагаемого материала и, одновременно, сопутствующая этому сложность изложения, обойти которую, несмотря на все старания так и не удалось, заставляют автора с извинениями рекомендовать при просмотре и чтении материала (конечно заинтересованным читателем), максимальное внимание. Запасшись терпением, не просто рассматривать рисунок как рекламированную ранее забавную картинку, но постараться тщательно разобраться в нем и понять вложенную в него сущность. Именно поэтому мы привлекли Ваше внимание к метаморфозе Эшера. Но ведь она велика еще и как творение Человека, хранит много скрытых тайн и достойна специального изучения. И почему - тоже нужно понять. Говорим мы об этом впервые! Автор постарается сделать все, что в его силах, чтобы облегчить эту, к сожалению, весьма нелегкую задачу. Автор |
Введение.
Структура и характер процесса трансформации клеточного множества биологического субстрата в процессе его озлокачествления претерпевает ряд структурных преобразований. Это – метаморфоза нормального эпителия в озлокачествленный. В процессе таких преобразований, реализуется специальная клеточная программа, реализация которой может быть отражена частотными распределениями отношения площадей клеточного ядра к площади всей клетки (ядерно - цитоплазматический индекс - ЯЦИ). Наиболее информативно распределение частостей, отражающих моду распределения этого отношения для совокупностей клеток основных фиксируемых состояний субстрата – норма, инициализация, предрак (дисплазия), рак. Вкратце мы говорили об этом в предыдущей статье, достаточно подробное изложение результатов вероятностных исследований клеточного субстрата изложено в статье . "Клеточный феномен "0.3-0.8"
http://science.pravda.ru/science/2003/6/18/50/15568_KLETFENOMEN.html
Динамика вероятностных отношений основных состояний субстрата совершенно неожиданно оказалось коррелированной с системой ситуативных отношений на графических изображениях - метаморфозах, созданных выдающимся голландским художником Мариусом Корнелисом Эшера. Соответствия метаморфоз - произведений изобразительного искусства и преобразований в живой природе не просто удивительны и интересны. Они позволяют глубже познать всю сложность скрытных отношений преобразований, связанных с отношением порождения – скачкообразным переходом от накопленной количественной определенности в новую объективную определенность предмета – его новое качество. Они также дают возможности открыть новую страницу в обсуждении творческих механизмов мозга, умственной деятельности художника, связывая ее с деятельностью рукотворной.
Маурис Корнелис Эшер (Maurits Cornelis Escher, 1898-1972) известен как мастер графического представления отношений между пространственными структурами, пространством и плоскими поверхностями. В большой серии своих работ, названной "метаморфозами" (1937-1945), он отобразил "забавные" графические структуры, казалось бы, не имеющие отношения к глубинам естествознания. Однако эти отображения, как в магическом зеркале, неумолимо подчиняются строгим формальным определениям, отражая самое существо динамики и кинетики естественных преобразований клеточных структур. Сегодня их можно считать аллегорическим отображением самой сущности, внутреннего содержания отношений преобразований в клеточных системах.
К сожалению, ни биоматематики, ни художественные критики, отмечая в свое время, динамичность и гармонию метаморфоз, полагая даже что они вносят музыкальный строй в изобразительное искусство, по достоинству не оценили этой замечательной и главной - познавательной особенности его творчества. Но они и не могли этого сделать - математика отношений в то время еще не была развита. Однако критики все же не могли не заметить в метаморфозах сам факт существования отношений как структурной компоненты художественного произведения [1], хотя и отнесли его значимость в ряду оценок особенностей произведений Эшера на последнее, 11-е место! Но сегодня мы в состоянии не просто содержательно интерпретировать эту связь, но более того, использовать метаморфозы Эшера как единственную реальную возможность, инструмент опосредования для конструктивного рассмотрения самой сущности сложнейшей феноменологии роста злокачественной опухоли, потери системой живого организма гомеостазического качества – сохранения жизнеспособности и замены этого качества сохранения обратным ему качеством антигомеостазиса - сохранения разрушения, "локальным гомеостазисом".
В настоящей статье производятся сопоставления и проводятся параллели, быть может спорные, но позволяющие выявить, обсудить и понять сложнейший мир отношений метаморфозы биологического субстрата путем анализа отношений на ее аллегорическом отображении – "Метаморфозе" Эшера.
Метаморфозы Эшера: одинаковость, сходство, порядок.
"Небо и вода" - так называется одно из произведений художника. "Метаморфоза" - другое. Что между ними общего и что их отличает? Что определяет общие особенности метаморфоз Эшера?

Рис. 1. М.К. Эшер "Небо и вода"
Не составляет особого труда заметить два основных объекта первого из них: это существа, обитающие в небе и воде - птица и рыба. Не доверяя внешней простоте рисунка и рассматривая изображения существ внимательно, можно увидеть и сущность метаморфозы: при движении по рисунку снизу вверх, накапливаются несущественные, едва различимые отличия у сходных объектов – наращивается количественная определенность.
Это приводит к новому объекту, совершенно не похожему на исходный. Малые накопления количества приводят к появлению новой объективной определенности - от рыбы к птице, В других метаморфозах Эшера одновременно генерируется и обратный переход – отображение динамического перерождения одного предмета в другой. Новое качество есть результат накопления малых количественных изменений, которые видны в промежутках между фигурками - на фоне плавного перехода от воды к небу.
Если же двигаться вдоль уровней изображений, мы видим их полное сходство: говорят, что объекты находятся в отношении одинаковости. Если для объектов указывается только сходство, то их невозможно разделить, сгруппировать на какие-либо четкие классы таким образом, что внутри класса объекты похожи, а вне его, между объектами различных классов сходства нет: говорят, что объекты находятся в отношении сходства. Отношение сходства имеет место в вертикальном сечении рисунка.
Оба сечения объединяет отношение упорядоченности или порядка. Отношение порядка соотносит элементы множества по выбранному признаку - важности, взаимному старшинству, первичности, геометрическому подобию и т.п. Признак упорядочения по вертикальному сечению метаморфозы - статичность градиента изменений графического подобия. В горизонтальном сечении упорядоченность образуется на множествах перемежающихся элементов, связанных отношением одинаковости и это также порядок.

Рис. 2. М.К. Эшер. Метаморфоза 1 (Methamorphosis) (1937)
На втором изображении (1937) ситуация существенно иная. Здесь художник передал свое понимание отношений для метаморфозы множества различных статических объектов (в данном случае - это множество архитектурных изображений местечка Амалфи на юге Италии) в единственный динамичный объект - забавного человечка. Метаморфоза забавна. Но в ней скрыт громадный смысл, она интерпретируя естественный эволюционный технологический процесс, отражает множество сложнейших ситуативных проблем и отношений на них.
Отношение одинаковости на ней сохраняется. Но оно "размыто", возникает и исчезает, скрытно перемежаясь с отношением сходства. Существуя в динамике трансформации, оно делает отношение сходства также размытым, признаки сходства незаметно переходят, перетекают друг в друга. Это отражает связь признаков одинаковости отношением динамичности, перетока. Как бы исчезая в одном множестве, они порождаются одновременно внутри другого деформирующегося множества. Проявляясь аналогичным образом в обеих (горизонтальном и вертикальном) сечениях, отношения одинаковости и сходства не просто нечетки, размыты, но и динамичны. Такой характер отношения одинаковости изменяет самую его сущность: от сходства в вертикальном и горизонтальном сечениях "нерегулярных" архитектурных и "регулярных" геометрических фигур – элементов метаморфозы, до сходства в горизонтальном и одинаковости в вертикальном сечении элементов множества, представляющих человеческие фигуры, а затем - их почти неразличимого сходства с фигурой живого существа – человечка.
Эти изменения отражают кинетический процесс трансформации не только отдельных элементов рисунка и их множеств, но и формирования некоторого динамичного множества подмножеств, связанных отношением перетока, сходных между собой, или более того, находящихся в отношении одинаковости. Это сущность отображения специфического механизма активации метаморфозы (внесенного творческим мышлением художника), соответствующая "вложенной" кинетике процесса изменения смысла отношений одинаковости и сходства, внутренней сущности компонент динамичности - "размывания", "сгущения" и "перетока".
Отвлекаясь от предмета обсуждения заметим, что если, с другой стороны, говорить о динамичности смысла этих компонент как отношений технологии отображения творческого замысла, то это - материализованное отражение конкретного творческого мышления как технологии "понимания", интеллекта художника и это дает возможность говорить о мышлении как о способе, технологии мозговой деятельности, например - обработки информации.
Динамичности отношений сходства и одинаковости соответствует также динамичное отношение упорядоченности. Но здесь отношение динамичности меняет свое качество - оно становится отражающим, рефлексивным. Изменяя смысл, оно разъясняет (эксплицирует) кинетические, "фронтальные" характеристики ветвления процесса трансформации. Первая же метаморфоза кинетических изменений не отражает, упорядочение носит монотонный однонаправленный характер. Можно говорить о том, что динамичности отношения упорядоченности свойственно скрытно отражать некие характерные внутренние динамические механизмы, порождающие развитие собственно процесса трансформации, хотя в изображение на рисунках они (как и другие отношения), естественно, внесены автором. Независимо от их происхождения, на метаморфозе можно зафиксировать одновременное существование обеих процессов - активации и трансформации. Это придает динамичности соревновательный характер, а отношениям сходства и одинаковости ситуативный смысл. Именно такого рода отношения существуют в динамике изменения состояния клеточного поля – метаморфозе биологического субстрата.
Изучением математических закономерностей статических отношений равенства, сходства и порядка занимался известный советский ученый математик-алгебраист Юлий Анатольевич Шрейдер. В 1971г. он издал книгу под названием "Равенство, сходство, порядок" [2]. Наши исследования динамичности ситуативных отношений метаморфоз биологического субстрата базируются именно на эту работу.
Отношения на сходствах.
Каждый объект в метаморфозе Эшера - элемент множества. Он несет информацию о похожих на него элементах, но эта информация отлична от той, которая содержится в случае отношения одинаковости. Здесь возможна различная степень информации, содержащейся в одном элементе относительно другого. Превосходная степень отношения сходства - неразличимость. Одинаковость связана с тем, что объекты неразличимые, как и сходные, нельзя разбить на классы таким образом, чтобы в каждом из классов элементы не различались, а разных классов различались заведомо. Отсюда следует, что одинаковость есть частный случай сходства. Понятие "одинаковость" также расплывчато, его точный смысл связан с учетом разъясняющих (эксплицитных) отношений эквивалентности и толерантности, которые применительно к метаморфозе клеточного материала рассматриваются специально. Размыты и ситуативные отношения - метаморфоза Эшера отражает множество размытых подситуаций без четких границ на геометрических элементах, образующих такие же размытые ситуации.
Но естественная динамичность размытости отношений, описания элементов множеств и ситуативности отношений на них, свойственна и любому отображению распределенных в пространстве биологического субстрата клеточных сообществ. Именно это и вносит существенные сложности в распознавание, описание и оценку его текущих состояний, в особенности - на ранних стадиях возникновения и развития процесса озлокачествления метаморфозы.
Постановка задачи отображения динамичности и размытости ситуативных отношений метаморфозы биологического субстрата, соответствует задаче отображения естественного технологического процесса потери его гомеостазического качества - сохранения жизнеспособности.. Каждая клетка из подмножеств их функционально дифференцированного множества, изменяясь, последовательно проходит через различные стадии озлокачествления, "перетекая" из одного подмножества озлокачествленных клеток в другое, функционально взаимодействуя при этом с клетками как своего, так и иных подмножеств, пока не закончит свою жизнедеятельность в одном из подмножеств клеток канонического состояния, возможно - в виде "голого" ядра множества клеток "раковой жемчужины", т. е. клетки отношение площади ядра которой к площади клетки, равно единице. Наблюдаемое в клинике изображение каждого элемента клеточного поля, как и поле в целом, обладает всеми необходимыми и достаточными признаками динамической размытости.
Как в произведении искусства отразить, интерпретировать и как опосредовать закономерности размывания и перетока отношений, характер ситуативности такого биологического феномена? Они ведь нам к тому же и неизвестны! Достаточно ли для этого аллегории художника, даже если это магическое зеркало Великого Эшера? Ответить на этот вопрос не легко. Но мы попытались это сделать, прибегая к помощи математики.
Будем обсуждать изображение и интерпретировать элементы образа метаморфозы в терминах естественной технологии размножающихся клеточных множеств, полагая, что каждый из интерпретируемых на картине элементов множеств отражает клетку, обладающую теми или иными функциональными, дифференцированными особенностями. Это приведет нас к обсуждению ситуативных отношений на метаморфозе, отражающих динамические отношения процессов трансформации биологического субстрата на всех компонентах множества - составляющих его подмножеств и отношениях на них. Так как в этих подмножествах, в свою очередь, отражаются функциональные группировки клеток, то тем самым отражаются и механизмы ситуативных отношений групповых взаимодействий между ними. Этим отражениям должны и могут быть поставлены в соответствие некоторые численные параметры - характеристики порождающих эти отношения биологических механизмов. Но если существуют характеристики отношений и их интерпретация, то существуют и могут быть определены возможности их использования!
Задачу феноменологического исследования естественного технологического процесса роста опухоли – механизма роста, можно формулировать как задачу множественного описания, численного представления и отображения множеств ситуативных отношений межклеточных функциональных взаимодействий в динамике трансформации биологического субстрата из нормального состояния в состояние озлокачествления, с учетом отношений кинетики текущих состояний субстрата на всем множестве механизмов групповых ситуативных отношений взаимодействий.
Естественно-технологический процесс озлокачествления и метаморфоза Эшера находятся в отношении ситуативного сходства. Это – сходство динамичности и оно определяется подобием перетоков клеточных ситуаций на решетке клеточных множеств и расплывчатых перетоков ситуаций на множествах графических элементов метаморфозы. В частности, достижение канонического состояния, представленного изолированным множеством голых ядер, находится в отношении сходства с формированием одиночной и изолированной, полностью сформированной фигурки человечка на метаморфозе. Она образовалась скачком, "оторвавшись" и освободившись от размытых ситуативных отношений метаморфозы, скачкообразно получив совершенно четкий "статус"! Динамика финала процесса озлокачествления имеет особенность именно такого рода: преобразование множеств клеток, на его завершающей стадии, в множество голых, совершенно "четких" ядер, происходит скачком, взрывообразно. Еще в 1973 году это статистическое явление роста отмечалось как феномен "ракового взрыва", завершающий естественный технологический патологический процесс [3].
Именно эта особенность метаморфозы Эшера натолкнула нас на мысль о ее сходстве с метаморфозой преобразования биологического субстрата.
Метаморфоза Эшера – аллегорический образ метаморфозы биологического субстрата.
Метаморфозу Эшера можно рассматривать по разному. Можно полагать ее просто забавным рисунком, показывающим, как из блока архитектурных сооружений путем некоторых ухищрений – приемов, уловок, или даже хитростей, получить фигурку человека. В этом случае можно установить связь между ее образом и смыслом и между ее значением и образом, что не требует внешней интерпретации. Это составляет ее изобразительную ценность – адекватно отразить видимое. Но ту же метаморфозу можно рассматривать с других позиций: можно полагать, что это - изображение отвлеченной идеи посредством образа и считать, что видимый образ отделен от смысла и связь между ее значением и образом устанавливается по сходству. В этом ее познавательная ценность – познать то, что в изображении скрытно. Сопоставляя какой либо трансформационный процесс с метаморфозой как с неким эталоном, можно найти, с некоторой степенью конкретизации, скрытные связи образа и смысла сопоставляемого процесса, его значения и образа. В этом случае метаморфоза рассматривается уже как аллегорическое отображение процесса трансформации и требует внешней интерпретации. В этом ее двойной, дуальный смысл.
Наша интерпретация метаморфозы, в предположении отношения ситуативного сходства сопоставляемых метаморфоз допускает, что она отображает множество размытых ситуативных отношений на множестве трансформирующихся элементов, воображаемых художником, и что это отображение, по крайней мере визуально, находится в отношении сходства с ситуативными отношениями на множествах преобразующихся элементов биологического субстрата. Однако одного внешнего сходства для утверждения такого предположения еще недостаточно. Для этого необходимо, чтобы оно, это сходство, было подтверждено на численной мере относительно численных критериев. Для такого подтверждения установим некоторые отношения сходства процесса озлокачествления и метаморфозы Эшера, путем сопоставления ключевых сечений образа метаморфозы и параметров частотных распределений значений ядерно-цитоплазматического отношения.
При большом количестве измерений частота появления тех или иных значений ядерно-цитоплазматического отношения в клетке, приближается к вероятности. Изменяясь непрерывно или дискретно от нуля до единицы, эти величины образуют вероятностную меру. Значения x этого отношения могут лежать в пределах от минимального, равного приблизительно 0.05, до значения равного единице для "голого" ядра - крайней степени озлокачествления клетки. Мы имеем возможность провести оценку сходства метаморфоз, используя вероятностную меру в пространстве ее существования.
Констатация такого сходства позволит полагать, что процесс последовательного преобразования одного частотного распределения ядерно-цитоплазматического отношения в другое, является отражением ситуативных отношений динамики трансформации элементов множеств и это означает существование сходства закономерностей в отображении обеих метаморфоз при их различном образе. При этом, конечно, нельзя забывать, что динамика трансформации субстрата порождается естественным путем - биологическими, внутриклеточными механизмами и межклеточными взаимодействиями клеточных множеств, в то время как метаморфозы Эшера продукт хотя и естественно-технологической, но сознательной рукотворной деятельности мозга.
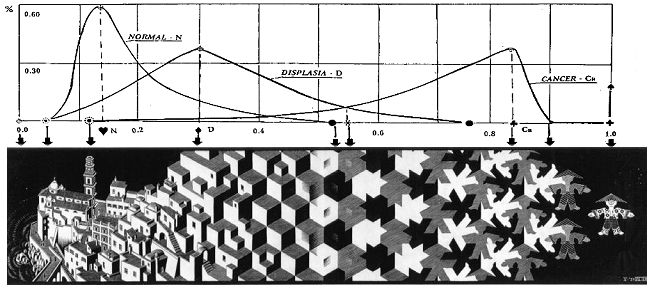
Рис. 3. Сопоставление положения и позиции
характерных точек частотных распределений и специфических сечений метаморфозы.
Первый шаг, который необходимо сделать для сопоставительного анализа, это убедиться, соответствует ли вероятностное описание текущих состояний субстрата и соответственно, его транспорта, переходов из состояния в состояние, множеству ситуативных отношений элементов метаморфозы Эшера. Одновременно нужно убедиться в наличии и обратного соответствия. Иначе говоря, необходимо взаимно сопоставить кривые распределений и ситуативные отношения на множествах элементов метаморфоз. Для этого проведем оценку взаимного положения, позиций характерных точек частотных распределений и сечений метаморфозы, соответствующих специфическим, размытым или четким отношениям ее элементов, множеств и свойственных этим множествам особенностей, например - в раскраске.
Будем отличать понятие “рассечения” метаморфозы, приводящего к получению наклонного или косого изображения в плоскости аппликаты пространственной системы координат и сечение изображения на две части плоскостью, перпендикулярной плоскости рисунка и разделяющее изображение на две части. Позиции характерных точек и сечений маркируются условными метками.
Вначале зафиксируем исходное состояние. Пусть ему соответствует четкое положение, обозначающее край скалы на левой границе очертаний набережной городка, строения которого подвергаются трансформации (рис.3). Это - "четкий" нуль отсчета в системе координат X0Z. Концу процесса логично противопоставить сечение оси симметрии (точнее - геометрического центра) полностью сформированной фигурки человечка - индикатора финитности, завершенности процесса. Это - тоже четкая фиксация, единица, и это образует четкий единичный интервал [0÷1]. Разобьем его на десять равных частей и оцифруем их. Этим создается единая система отсчета, соответствующая отсчету значений ядерно-цитоплазматического отношения и относительная мера на ней, позволяющие реализовать сопоставительный анализ частотных распределений клеток различных состояний, с ситуациями на метаморфозе, образуемыми различными (четкими и размытыми) сечениями (четких и размытых) множеств таких же (четких и размытых) изображений элементов метаморфозы, их (четких и размытых) объединений, пересечений и границ. Оба объекта сравнения обладают аналогичными свойствами, однако в метаморфозе Эшера эти свойства, формируя образ видимы, а в метаморфозе биологического субстрата, представленной своими распределениями, они скрыты. Поэтому при сопоставлении метаморфоза выступает в двух ролях – с одной стороны как произведение искусства, составляющее изобразительную ценность, с другой – как аллегория, в которой образ отделен от смысла и который может быть ему придан (путем внешней интерпретации) по сходству с метаморфозой биологического субстрата.
Согласовав совместное положение начальных и конечных точек отсчета распределений и картины метаморфозы, имеющих теперь одинаковый линейный масштаб и общий отсчет положения элементов, рассмотрим ситуативные отношения сходства и одинаковости на элементах ее множества в сечениях, соответствующих значениям мод распределений – констант состояния, пересечениям распределений и другим характерным значениям x признаков озлокачествления и особенностям распределений.
Двигаясь от начала отсчета, начиная с вполне четких изображений элементов архитектуры, не трудно увидеть (на самом деле для такого видения необходимо максимальное сосредоточение внимания!), что появление первых дисплазионных клеток ž совпадает с инициацией появления некоторого множества ее черных цветовых элементов. Положению ♥ максимума плотности вероятности нормального эпителия (мода N), при одновременном появлении на кривой D ~13% дисплазионных клеток, соответствует вертикальное сечение на множестве архитектурных элементов изображения, которое можно охарактеризовать как сечение активации "размывания": в этом сечении зримо начинают закрываться не подвергающиеся в дальнейшем размыванию архитектурные сооружения заднего плана рисунка. Одновременно, на его переднем плане (и это подчеркивает лидирующую в процессе трансформации роль динамизма отношений), активизируется процесс преобразования двухцветных (серо-белых) объемных очертаний архитектурных образов в размытые регулярные геометрические трехцветные объемные фигуры. Распространение процесса такой активации направлено по оси ординат (вверх) и в глубину рассечения изображения (горизонтальные плоскости объемных элементов метаморфозы, окрашенные серым цветом, параллельны основанию, на котором возведены сооружения). При этом, в сечении, соответствующем проекции пересечения кривых распределений N и D (ядерно-цитоплазматическое отношение ~0.225), задний план, не подвергающейся трансформации архитектуры, полностью закрывается элементами переднего плана и исчезает.
Это отражает приобретение множествами элементов метаморфозы не только нового качества, но и новых функциональных отношений, отображенных в новые группировки ее элементов. Отдельные поверхности компонент архитектуры черной окраски, появившиеся на верхних и правых, боковых поверхностях элементов нижней части этого же сечения, отображают формирование некоторого их подмножества - активаторов, порождающих и развивающих направление распространения процесса активации в направлении оси отсчета (количество таких подмножеств на метаморфозе совпадает с числом подмножеств клеток - активаторов процесса, фактически отмеченных по данным клиники на гистологических препаратах). По мере перемещения вправо, изображения архитектурных элементов размываются, образуя изображения геометрических объемных фигур, принимающих все более четкое очертание. Размывание архитектурных изображений и дочерних, порождаемых ими геометрических фигур, захватывая весь объем пространства трансформации, продолжается вначале как трехмерный объемный процесс, вырождаясь затем в двумерный, плоскостной.
Значимый момент: здесь появляется своя, независимая трехмерная система координат X’0’Y’Z’с началом в элементе черной окраски (см. внимательно, сравнивая с рис.4!). Этот элемент отображает инициальное подмножество клеток - активаторов процесса и это отражает образование трехмерного пространства процесса активации. Начало 0’ этой системы координат лежит точно в сечении ž начала появления озлокачествленных клеток распределения Ca – рака (и, одновременно, в близости слева к сечению ♥ моды распределения N нормального эпителия). Эта точка отображает относительный временной параметр начала процесса инициализации. На интервале [♥,u] между модами распределений N и D объемная трансформация архитектурных элементов метаморфозы размывается (отношения сходства теряются) во всех трех измерениях. Начиная со значения u (~0.3) ядерно-цитоплазматического отношения, в непосредственной близости к константе перехода от пластического в гиперпластическое состояние, размывание вдоль вертикального сечения постепенно увеличивается и замещается отношением объемного сходства, переходящего в отношение плоскостной одинаковости элементов по вертикальным сечениям вблизи (слева) от точки · затухания распределения нормального эпителия. Далее, в направлении индикатора "финитности", отношение сходства последовательно по сечениям размыто перерастает в отношение одинаковости.
Однако, если сопоставляемые метаморфозы сами находятся в отношении сходства, то аналогичные динамизмы отношений сходства и одинаковости их элементов, свойственны и метаморфозе биологического субстрата. А это означает, что параметры состояния субстрата, перетекая из множества параметров эквивалентности (отражающих сходство), приобретают свойства толерантности параметров (отражающих одинаковость). Свойство толерантности свойственно параметрам субстрата, отражающим переход клеток в каноническое, озлокачествленное состояние.
Появление трехмерной системы координат в точке 0’, сдвинутой и развернутой в пространстве рисунка относительно исходной плоской системы отсчета (рис.4), отражает не просто значимый, но этапный момент в развитии процесса. Оно отображает появление новой и направленной степени свободы - пространства размножения клеточных популяций, начало процесса инициализации.
Трем мерностям пространства соответствуют три цвета раскраски граней изображений элементов метаморфозы Эшера: белая окраска соответствуют левой боковой, серая – горизонтальной, параллельной основанию объемной фигуры изображения, а черная – правой боковой их поверхности. Можно представить, что множество элементов изображения, кажущихся плоскостными после момента x=0.5, но существовавших до этого в трехмерном пространстве, потеряв одну мерность, оказывается как бы "сваленным" на одну из своих граней. Это можно расценить как отображение начала деформации образов таким образом, что по мере приближения к центру индикатора финитности – фигурке человечка, множество объемных элементов метаморфозы "сжимается" в множество плоских элементов. Четкое отображение отношения деформируемости в сечении · метаморфозы, четко соответствует конечному значению ядерно-цитоплазматического отношения для нормального эпителия и находится в близости слева к точке Ú обращения эпителия в отношение статистической "левизны-правизны" распределений D и Cа. Этот факт также отражает этапный момент процесса. Цветовая размытость, соответствующая объемно-плоскостным преобразованиям элементов метаморфозы, весьма четко отображает на интервале [·,Ú] переход к плоским изображениям в общем направлении индикатора финитности.
Из этого логически следует, что в третьем измерении процесса трансформации – плоскости рассечения изображения, отражается существование, в динамике перетоков в каноническое состояние, некоторого подмножества молодых клеток нормального состояния N (это, как показало исследование, отражается и на «решетке» множеств – их пересечениях и объединениях). Исчерпание этого подмножества в процессе транспорта, при значении x близком слева к x=0.5, отражается исчезновением мерности пространства в боковой плоскости.
Сжатие образа множеств элементов в плоскости изображения отражает направленность процесса в сторону индикатора финитности. Аналогично, правосторонняя направленность в динамике роста отображается и на решетке множеств, образующей характеристическую поверхность. Это - следствие отношения следования между физическими процессами, отражаемыми плотностью окраски ядра и ростом ядерно-цитоплазматического отношения. Сам же интервал [·,Ú] сжимаемости оказался нечетким, скорее всего из-за естественного накопления относительных погрешностей измерений и обработки малых значений ядерно-цитоплазматического отношения на конце распределения для нормального состояния эпителия. Обработка данных вносит свою размытость и свои сложности.
Вернемся к рассмотрению динамики преобразования на метаморфозе. Она отражается в отношения одинаковости не только по форме, геометрии элементов, но и по их окраске, охватывая полный комплекс признаков изображения отдельных элементов. Плоские фигуры на интервале [·,Ú] обладают отношениями одинаковости как по окраске, так и по геометрии в вертикальном сечении рисунка, а между их подмножествами образовались размытые отношения сходства. Затем размытые плоские фигуры, одновременно наращивая в каждом цвете отношения сходства по направлению финитности и по двум диагональным направлениям, а также одинаковости в вертикальных сечениях, преобразуются в фигурки человечка трех цветовых оттенков. При этом элементы множеств, определенные на интервале [·,Ú], могут рассматриваться, как центры порождения подмножеств элементов индикатора финитности. Черные и белые фигурки попарно направлены головами навстречу друг к другу под углом 120º и перемещаются параллельно в сторону роста по пересекающимся направлениям слева сверху - направо вниз и слева снизу – направо вверх. Таким образом "равнодействующая" активации и трансформации направлена точно в центр сформировавшейся финитной фигурки, однако черная фигурка движется к нему "головой" вперед, а белая – “ногами”. И это тоже имеет свое объяснение. Серая компонента финитной фигурки формируется без вращения (это может получить свою трактовку) и, перемещаясь по направлению равнодействующей, на интервале [♣-0.9], сливаясь с черной и белой компонентами, приобретает очертания индикатора финитности. Одновременно, на этом же интервале [♣-0.9] черные и белые фигурки человечка, находясь в отношении сходства, близком к одинаковости, разворачиваясь (на разные углы) вокруг своих центров (совпадающих для черных фигурок с положением ♣ раковой константы) и сливаясь с серой фигуркой, полностью формируют свой конечный, финитный облик. При "обнулении" функции распределения Ca исчезает четкая черно-серо-белая окраска, дифференцирующая подмножества фигурок, а затем в интервале [0.9-1.0] исчезают и сами фигурки, которые появляясь скачком, в сечении Ì принимают свой окончательный четкий, как по геометрии так и по окраске, двухцветный вид. На распределениях ядерно-цитоплазматического отношения в этом диапазоне, клетки вообще не наблюдаются (частость их появления равна нулю). В сечении Ì, соответствующем концу процесса трансформации и совпадающим с центром сформированной (четкой) финитной фигурки, образуется скачек ♠ в виде дельта-функции распределения ядерно-цитоплазматического отношения с мощностью множества голых ядер в 16%. Сформировавшееся полностью финитное изображение человечка в этом сечении – отображение ракового взрыва.
В расцветке элементов метаморфозы в начале процесса преобладает бело-серая окраска. Она размыто замещается черной окраской, интенсивность которой суммарно по площади рисунка возрастает с начала процесса трансформации. Серая окраска, имея слабо возрастающую плотность до момента полного затухания распределения нормального эпителия, в интервале сжимаемости [·,Ú] - порождения подмножеств элементов индикатора финитности, начинает размываться постепенно и равномерно, исчезая полностью к моменту формирования финитной фигуры. Динамизм изменения цветности отражает такие изменения элементов изображения, параметры которых, образуя множество параметров эквивалентности, перетекают в множество элементов, параметры которых обладают свойством толерантности.
Распространяя это высказывание на метаморфозу биологического субстрата, можно сказать, что белая окраска отображает рост опухоли, опережая текущее состояние, черная - текущее состояние субстрата, а серая соответствует отсчету, увы, "редуцированного" от момента окончания, время процесса.
Угловое положение формирующихся фигурок метаморфозы, как и различие в их окраске, имеют свою трактовку. Положение черных и белых фигурок связывается с отображением двунаправленного действия двух различных механизмов агрессии подмножеств клеток, функционально-ориентированных на активацию процесса разрастания опухоли. Существование таких механизмов отражается и на решетке множеств характеристического поля, и на многочисленных графах состояния субстрата.. На образе метаморфозы их взаимодействие отражает преобразования серых фигурок, которым можно приписать отображение прогрессирования процесса активации, направленного непосредственно в центр индикатора финитности. Углы наклона фигурок человечка отражают направление вектора равнодействующей изменения состояния субстрата, а формирование их комбинированной (черной, серой, белой и пятнистой) окраски, отражает изменение четырех градаций оптических показателей ядер клеток, аналогичных градациям, образующим систему отсчета значений прозрачности клеточного ядра при подсчете обобщенного показателя озлокачествления.
Сопоставительный анализ распределений ядерно-цитоплазматического отношения и ситуативных отношений на множествах элементов метаморфозы Эшера показывает, что сама возможность отображения процесса трансформации биологического субстрата существует, сущность задачи построения такого отображения однозначно иллюстрируется и формулируется благодаря дуальности свойств метаморфозы Эшера. И в этом – величайшая заслуга художника перед человечеством.
Теперь, независимо от предметности внешней интерпретации, абстрагируясь от образа, значения, сходства и каких либо аналогий, можно сформулировать сущность, выражающую единство многообразных свойств и отношений творения Эшера, как
- ситуацию на множестве подмножеств трансформирующихся элементов, находящихся в динамических отношениях сходства и одинаковости во времени и пространстве, стремящемся от четкости исходных описаний и отношений, через множество четко упорядочиваемых в процессе трансформации отношений размытости описаний и механизма формирования подмножеств, отражаемых множеством показателей эквивалентности, к четкому финитному отношению завершенности процесса, отраженному множеством показателей толерантности.
И главное в сущности метаморфозы – четкое динамическое упорядочение отношений размытости отображения механизмов, формирующих четкие связи между динамически изменяемыми размытыми множествами размытых элементов, участвующих в процессе. Такая философско-математическая сущность есть «внутренность» каждого множества элементов метаморфоз Эшера. Размытость отражает процессы медленных количественных накоплений порождения, скачкообразно переходящих в четкие качественные изменения.
Сделаем одно замечание, касающееся исходной структуры для гипотетического построения (не исключая такой возможности в принципе) метаморфозы биологического субстрата формальным, аналитическим путем. Метаморфоза Эшера отображает процесс трансформации цельного подмножества (блока) из множества архитектурных сооружений, "вырезанного" из городского пейзажа. Можно говорить о том, что аллегория Эшера, отправляясь от одиночного блока сооружений, может быть сопоставлена лишь с преобразованием одного функционального подмножества клеток биологического субстрата. Если бы художник задумал отобразить параллельно и одновременно метаморфозу для композиции групп строений, он для их дифференцировки, использовал бы кроме человечков иные индикаторы преобразований, например - мифические или другие любимые им образы. Но ситуативные отношения и динамика их преобразований не зависят ни от характера ни от мощности множества наборов первоначальных образов, а целевая функция преобразования такова, что независимо, также и от образа художественного воспроизведения в целом, трансформация всегда преследует цель достижения финитного состояния, реализуя эту цель едиными механизмами.
Это означает, что аналитическое построение метаморфозы по данным клиники биологического субстрата может учитывать, а может и не учитывать дифференциацию первоначальных наборов клеток и соответствует утверждению, что в процессе трансформации они не теряют свою первоначальную функциональную дифференцировку, а изменяют ее, причем в динамике трансформации - неоднократно.
В метаморфозе, построенной аналитическим путем, может быть, а может и не быть ни аллегорических образов, ни каких либо художественных изображений, однако ее представление в виде геометрических фигур, может иметь достаточно сложные формы. Как и другие математические построения, ее архитектурные элементы и отношения на них, должны получить строгую внешнюю интерпретацию.
Динамизмы на метаморфозах.
Можно считать, что метаморфоза трансформации биологического субстрата, в том числе и отраженная аналитическим путем, имеет специфический статус информационного динамического преобразователя отношений сходства и одинаковости на визуальной информации (образе), отраженных на динамизмы преобразования множества эквивалентности значений показателей озлокачествления в множество их толерантности. Эти динамизмы можно проследить непосредственно на метаморфозе Эшера, путем сопоставления динамики изменяемости ее структуры с динамикой изменения ядерно-цитоплазматического отношения.
Как отмечалось, сопоставление метаморфоз показывает совпадение положений возникновения некоторого подмножества вполне черного цвета элементов архитектуры на переднем плане метаморфозы, с появлением первых клеток гиперпластического состояния Ca (Cancer) на распределениях ядерно-цитоплазматического отношения и, одновременно, с накоплением 13% клеток пластического состояния D (Displasia). Сечение такого тройного совпадения мы определили как сечение активации размывания изображений на метаморфозе и, соответственно ему, инициации процесса озлокачествления в биологическом субстрате. Это означает, что начало процесса инициализации соответствует (по существу?) началу "размывания" межклеточных отношений. Оно отражается возникновением, в сечении активации, пространственной системы координат X'0'Y'Z' (рис.4). В этой системе по оси ординат 0'X' отражаются изменения хроматических свойств трехцветного преобразования элементов изображения, а их распределение лежит в плоскости X'0'Y' рассечения.
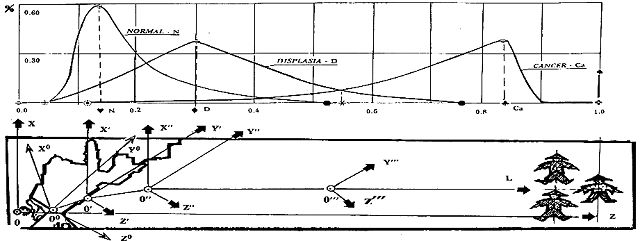
Рис. 4. Динамические соответствия на метаморфозах.
Пространство активации занимает интервал от начала 0' этой системы координат метаморфозы до сечения, соответствующего значению 0,225 пересечения распределений нормального и дисплазионного состояния.
В этом сечении, в точке, сдвинутой относительно начала координат 0' координатной системы X'0'Y'Z' на величину DZ' вдоль оси 0'Z' и на величину DY' вдоль оси 0'Y', образуется вторая система пространственных координат X''0''Y''Z''. В этой системе, биссектриса координатного угла 0''L координатной плоскости Y''0''Z'', параллельной плоскости основания изображения, оказывается направленной точно в центр фигуры - индикатора финитности. Сдвиг начала координат этой системы на величину DX=0.15, соответствует приращению ядерно-цитоплазматического отношения также на 0.15, точно совпадающему с константой нормального состояния и фиксирует на метаморфозе точку 0'', определенную по данным клиники как точку гомеостазического перехода в антагонистическое гомеостазису состояние озлокачествления, а положение биссектрисы координатного угла отражает вращение оси 0''Z'' на 45º против часовой стрелки. Это соответствует перемещению начала 0' системы координат X'0'Y'Z' в точку 0'' с одновременным ее вращением и образует, таким образом, вторую пространственную систему координат. Перемещение начала координат из положения 0' в положение 0'', предваряя возникновение состояния PRG0 на величину DX=0.075 ядерно-цитоплазматического отношения, заранее подготавливает условия для целевого управления динамизмами на биссектрисе координатного угла 0''L третьей системы координат. Все указанные геометрические показатели нашли свое отражение на математической модели процесса опухолеобразования.
В сечении изображения, соответствующем середине интервала сжимаемости [·,Ú], мерность X'' второй ситуативной системы координат исчезает. Это приводит к возникновению еще одной, но уже плоской системы координат Y'''0'''Z''', c той же биссектрисой координатного угла L, относительно которой изображения элементов оказались плоскими и развернутыми. Не исключено, что скачкообразная деформация второй пространственной системы координат, "сжимая" ее в плоскую систему Y'''0'''Z''', отражает переключение отношения "левизны-правизны" метаморфозы биологического субстрата, поскольку этот статистический феномен находится в окрестностях Ú сечения метаморфозы, близких к положению возникшей плоскостной системы. В пространстве параметров метаморфозы этому соответствует обращение множества эквивалентности параметров состояния в множество толерантности а на множестве отношений - обращение отношений размытости на отношения четкости.
При вращении системы X''0''Y''Z''образуется пространство, ограниченное координатными плоскостями Y'0'X', Y''0''X'', и X'0'Z', X''0''Z'', содержащее в своем "г" –образном объеме конечное подмножество элементов активации отношения размытости. Ему можно поставить в соответствие подмножество клеток баланса эквивалентности в интервале распределения ЯЦИ 0.13£X£0.25 и соответствующие подмножества клеток на решетке исходного описания канонического состояния. Можно полагать, что этому пространству на метаморфозе соответствует динамический переток изображений сквозь образовавшуюся поверхность X'0'0''X'' из области, ограниченной плоскостями X'0'Z', X''0''Z''в область, ограниченную плоскостями X''0''Y'', X'';0''Y''. На множестве клеток биологического субстрата, этому перетоку изображений соответствует клеточный транспорт множества инициированных клеток в множество клеток баланса эквивалентности. Поверхность такого транспортного перетока на множествах клеточных подмножеств, отражается как линейчатая, соответствующая характеру объема пространства активации на метаморфозе.
На множестве элементов метаморфозы, можно выделить еще одну пространственную область, ограниченную "начальной" системой координат X000Y0, относительно которой система X'0'Y'Z' оказывается сдвинутой на DZ0 и DY0 относительно соответствующих осей. Начало координат этой области лежит в точке, соответствующей проекции на изображение метаморфозы начала ž возникновения дисплазионных (и нормальных) клеток, а ее ось Y0 направлена с незначительным разворотом против часовой стрелки от системы X'0'Y'Z' в сторону, противоположную возникающему в начале координат 0' изменению цветности архитектурных сооружений.
Поверхность перетока X0000'X' метаморфозы рассматривается как инициальное дополнение участка поверхности перетока изображений X'0'0''X'' и в соответствие этой области сопоставляется область образования множества клеток состояния инициации подмножества клеток пластического состояния спокойного эпителия. Поверхность перетока получает незначительный, но важный стартовый разворот, порождающий отражение транспорта вдоль линейчатой поверхности перетока, обладающей криволинейностью, соответствующей изгибной линии конкатенирующихся биссектрис координатного угла последовательно порождаемых систем координат, как отражения целевого направления клеточного транспорта в финитную точку. Криволинейность поверхности перетока отражает конкатенацию объемов, образуемых двумя последовательными вращениями систем координат, как правостороннюю направленность решетки множеств на характеристическом поле, а ее образующей - как отражение стыковки поверхности перетока X'0'0''X'' и ее дополнения X0000'X'. Все изменения поверхностей перетока также нашли свое отображение на математической модели процесса.
Упорядоченная во времени последовательность появления систем отсчета на метаморфозе Эшера, находится в отношении соответствия ролевых функций множества подмножеств клеток биологического субстрата, связанных в свою очередь отношением порождения – перехода медленных количественных изменений в скачкообразные качественные. Четыре системы отсчета метаморфозы Эшера соответствуют числу существующих базовых систем ситуативных отношений метаморфозы биологического субстрата. Это системы гомеостазиса, гомеостазического переключения, клеточного транспорта в системе локального гомеостазиса и плоская система отображения финитных отношений на решетке множеств. Единство отображения в этих системах реализуется на мере сопоставимости обобщенного показателя озлокачествления, определенного на плоскостной масштабной системе отсчета.
Такие отображения, в виде наборов численных показателей, характеризующих процесс, идентифицируют вершины графа изменения состояний. Это устанавливает на нем отношение упорядочения причинно-следственных связей, аналогичное упорядоченности следования частотных распределений ядерно-цитоплазматического отношения, в том числе - в его сопоставительном анализе с образом метаморфозы Эшера.
В этой связи нельзя обойти вниманием один специфический вопрос, касающийся аналитического представления этой аллегорической метаморфозы. Характерные параметры динамики изменения вероятностных распределений ядерно-цитоплазматического отношения на метаморфозе биологического субстрата, совпадают с характерными сечениями образа метаморфозы Эшера. Не означает ли это, что существует численно определенный и обобщенный параметр образа элементов архитектуры, решетка множеств значений которого отражает динамику ситуативных отношений на множествах элементов этой аллегорической метаморфозы аналогично тому, как это имеет место в метаморфозе биологического субстрата? Иными словами: не является ли распределение Пирсона типа III (или гамма-распределение) тем же самым распределением, которое отражает частости появления некоего характерного, например - площадного показателя на множестве элементов образа метаморфозы Эшера, распределенного по его поверхности? Таким показателем в частности, могут быть отношения различной цветности площадей составляющих элементов, как и градации цветности, их окраски. И вообще, не является ли это распределение, в силу своих специфических особенностей (см. указанный сайт), статистическим отражением фундаментального отношения порождения, органически присущего феномену размножающихся множеств?
Мы не имеем доказательного ответа на этот вопрос, требующий проведения специальных исследований и специфической аппаратуры, но можем высказать предположение, логически вытекающее из сформулированных ранее результатов:
- да, существование такого обобщенного параметра, численно определяющего отношения на множестве элементов аллегорического образа метаморфозы, не исключено, и если он существует, он носит характер информации, содержащейся на ситуативных отношениях перетоков множеств ее элементов. Не исключено также, что количество этой информации находится в отношении соответствия с количеством информации, содержащейся на ситуативных отношениях перетоков множеств клеток биологического субстрата. Величина, характеризующая такое согласование, численно определяет сходство метаморфоз и это позволяет говорить о информационном характере клеточного антигомеостазиса.
Ниже мы сформулируем некоторые выводы, касающиеся параллелей между метаморфозами Эшера и феноменологией метаморфозы биологического субстрата. Эти выводы базируются на проведенный анализ. Нам представляется, что ознакомление с ними может породить интерес не только со стороны специалистов - биоматематиков, но и со стороны специалистов различного профиля, интересующихся прикладными проблемами "чистой" науки.
Заключение.
1.Множество трансформирующихся элементов метаморфозы биологического субстрата, находясь в динамических ситуативных отношениях во времени и в пространстве, стремится от нечеткости описания отношений инициализации клеточного субстрата, через множество упорядочиваемых в процессе трансформации отношений размытости механизма их формирования и описания элементов множеств, к четкому отношению строгого упорядочения финитности процесса.
2.Система динамических отношений на метаморфозе Эшера аллегорично отражает систему отношений процесса озлокачествления биологического субстрата. Естественная динамичность размытости отношений, описания элементов множеств и ситуативности отношений на них, свойственна любому отображению клеточных полей.
3. Отношение порождения на метаморфозе Эшера, динамически отображаясь в отношение соревновательности отношений сходства и одинаковости ее элементов, находится в отношении соответствия с феноменом обращения множества клеток эквивалентности биологического субстрата в множество клеток толерантности.
4. Метаморфоза Эшера, как аллегорический образ, может быть интерпретирована в терминах процесса опухолеобразования клеточных множеств, как отображение трехкомпонентных изменений ситуативных отношений на порядках изменения состояния клеточных множеств, их вторичного изменения как последствия активации и выбора направления активации, как равнодействующей изменения отношений между его активаторами.
Литература
1. Bruno Ernst. The Magic Mirror of M.C.Escher. Taschen America Inc. 1994
2. Шрейдер Ю.А. Равенство, cходство, порядок. Наука; М. 1974, cтр. 254.
3. W. Rоss Ashby The Set Theory of Mechanism and Homeostasis. "General Systems", vol. IX, 1964, p. 83-97.
4. Поспелов Д.А. Логико-лингвистические модели в системах управления. М. Энергоиздат, 1981, стр.230.
5. Поспелов Д.А. Ситуационное управление. Теория и практика. М. Наука, 1986б стр. 285.
6. Гельфандбейн Я. А., Каплан Б. Л., Методы статистической идентификации в задачах исследования и диагностики малигнизированных структур. Материалы II Всесоюзного симпозиума по биокибернетике. М. 1974.
7. Гельфандбейн Я А., Гельфандбейн В. Я. Проблема классификации и множественное описание размножающихся популяций.//АВТ, 1995, - № 6, - с. 68 - 81.
8. Гельфандбейн Я. А. Гельфандбейн В. Я. Аксиоматическая модель характеристического поля карциногенеза //АВТ.-1995. -№5.- c 80-84.
9. Ya.Gelfandbein, V.Gelfandbein. Classes of states and characteristic function of epithelial field malignization process. Automatic Control and Computer Sciences, N.Y., vol,28, No.5, pp.31-37, 1994.
10. Ya.Gelfandbein, V.Gelfandbein. The Classification problem and a multiple description of proliferating populations. Automatic Control and Computer Sciences, N.Y. vol,29, No.6, pp.68-81, 1995.
11. Ya.Gelfandbein, V.Gelfandbein. An Axiomatic model of the characteristic field of carcinogenesis. Automatic Control and Computer Sciences, N.Y. vol,28, No.5, pp.31-37, 1994.